Why are sensory attacks different to motor attacks? #MSBlog #MSResearch
“When we profile MSers, initial symptoms and the nature of early relapses have prognostic significance. It is generally accepted that sensory and visual symptoms are associated with a better outcome than motor attacks. Why? Recently when I was in Australia I presented my theory of MS being a length-dependent central axonopathy (disease of axons), which as you know is based on my observations of therapeutic lag and my asynchronous progressive MS hypothesis. Although I got a lot of nodding heads in the audience, not all agreed with my hypotheses. The work of Tomas Kalincik and others at MSBase (see below) was quoted as a possible exception to the rule. How does the length-dependent hypothesis explain the differential presentation of early and late attacks and their differential prognosis? Sensory relapses were more frequent in early or non-progressive disease. In comparison motor, sphincter (bladder and bowel) and cerebellar (incoordination) relapses were more common in older MSers and in progressive disease. Sensory, visual and brainstem relapses also showed better recovery than other relapses."
"Since my presentation in Melbourne I have been thinking about little else. It is well known that dirty little facts slay beautiful hypotheses. I may have a preliminary explanation based on the architecture and design of the nervous system to explain some of these observations. I have broken up the problem into several components:
1. Why are sensory attacks more common early on in the course of the disease?
The sensory system is synonymous with cutaneous sensation (light touch, pain, temperature, deep pressure, tickle, etc.). However, sensory should refer to all senses and hence should include vision, hearing, joint & head position, taste and smell. Physiologists call these the afferent systems; afferent simply means a neuronal input into the nervous system. In comparison, motor or output systems are called efferent systems. Afferent systems are designed for sensitivity; they need to pick-up and sense our environment so as to warn of us danger so that we can respond to them. From an evolutionary perspective our afferent/sensory systems are optimised for detection. A very large part of our cortex is devoted to perceiving the senses. In fact most of the parietal, occipital and temporal lobes are devoted to perceiving, integrating and interpreting sensory inputs. In comparison, the efferent/motor system is optimised for action and much less of our cerebral cortex is dedicated to motor activity. As a result of its size (cortical area) and optimised sensitivity a small lesion in the sensory pathways may be more likely to cause symptoms than a similar sized lesion in a motor pathway. This may explain why motor attacks have a worse outcome; larger lesions may be associated with greater damage.
3. Could reserve capacity explain the different prognostic significance of sensory and motor attacks?
Extending the arguments above that the sensory system may be more sensitive to small lesions due the reporting of percepts, in particular positive phenomena, that as it degenerates, loses neurones and axons, the likelihood of new lesions causing symptoms, in particular positive symptoms, diminishes. In comparison, as the motor system degenerates and compensatory mechanism fail new lesions are more likely to become clinically apparent. If reserve capacity in a particular pathway determines its sensitivity to cause relapses then one could hypothesise that proportion of positive sensory attacks (positive sensory symptoms) will diminish, in comparison to negative sensory attacks (loss of sensation). The latter hypothesis is testable and could be explored in existing databases or prospectively in clinical cohorts."
In my length-dependent model I propose that MS lesions may affect the nervous system randomly. Therefore, random MS lesions are more likely to hit sensory or afferent pathways, than motor pathways. We also have to get away from the dogma that MS is a white matter disease that only affects the so called subcortical white matter pathways. Recent pathological studies indicate that at least half the MS disease burden is found in cortical and subcortical gray matter. A small gray matter lesion in the sensory cortex could be a very common cause of early attacks. In comparison, a small lesion in the motor cortex may be less likely to cause symptoms. Please remember the sensory cortex is designed for perception that triggers awareness of our environment. In comparison the motor system is activated in via reflex, or through conscious volition. The motor system triggers a percept via a sensory feedback loops; for example we only perceive weakness because the sensory receptors in the joints and muscles tell our brains that there is mismatch between what is meant to be happening and what is happening. The feedback loop may result in the motor system having a much higher threshold; hence small or relatively minor lesions in the motor system may go unnoticed.
 |
| The sensory fibre tracts |
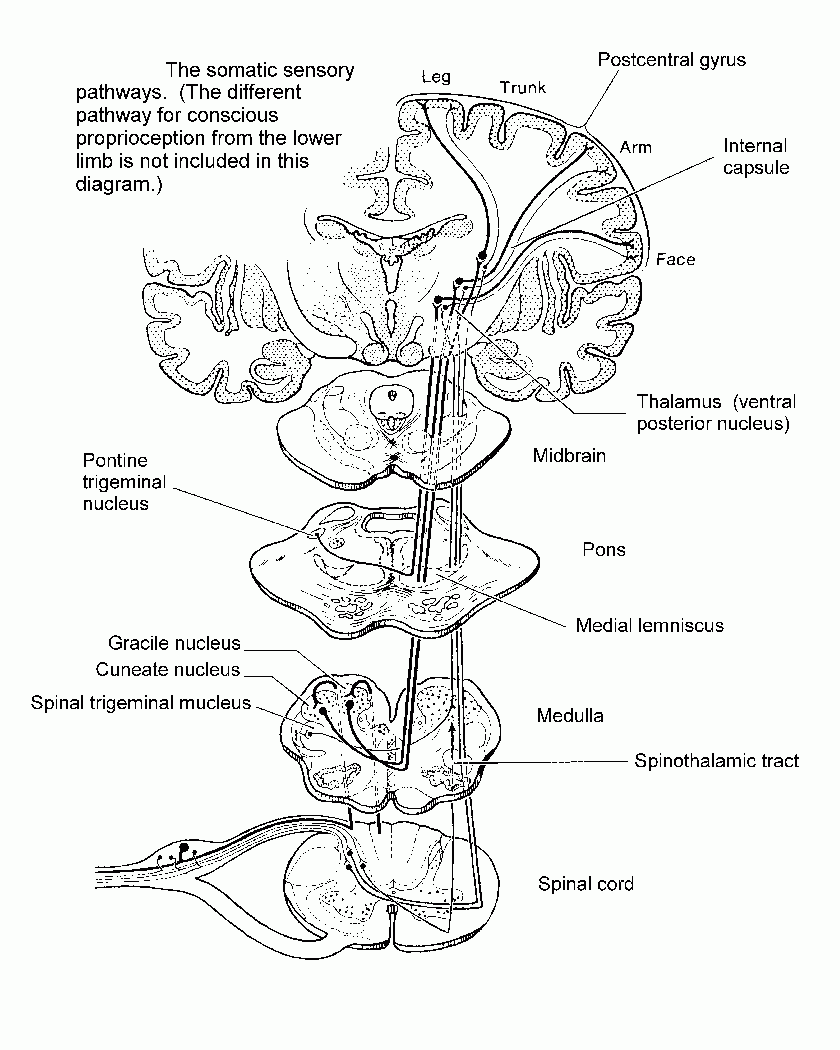{kind=link}
2. Could length-dependency explain the different prognostic significance of sensory and motor attacks?
The sensory system has shorter axons, compared to the motor system, and according to the length-dependent axonopathy hypothesis this may protect it. In comparison, motor neurones are much longer than sensory neurones and hence more likely to acquire multiple hits that lead to more rapid or earlier neurodegeneration. The architecture of the sensory system is such that afferent inputs go via first, second and third order neurones that communicate via synapses before reaching the cortex. Each relay axon is relatively short and hence this architecture may provide the sensory system relative protection from MS-associated neurodegeneration. In fact, the first-order sensory neurones are actually bipolar cells with their cell bodies residing outside the central nervous system in the dorsal, or posterior, root ganglia (DRG). A demyelinating plaque affecting the central axonal process is therefore unlikely to kill the neuron residing in the DRG by retrograde neurodegeneration. We know this because sensory radiculopathies that destroy the central neuronal processes before they enter the spinal cord are characterised by preservation of sensory nerve action potentials (SNAPs), which indicates that the neuronal cell bodies and peripheral axons are intact. The compartmentalisation of 1st-order sensory neuronal cell bodies, outside the CNS, away from the MS pathology, may provide the sensory pathway with a better chance of recovery, via axonal sprouting or axonal regrowth.
In comparison the axons from the pyramidal, or motor, neurons extend from the primary motor cortex to synapse on the anterior horn cells, or their equivalents, in the lateral columns of the spinal cord or brain stem respectively. Importantly, there are no motor interneurons, which is why the motor neurones are the longest neurones in the central nervous system. The cell bodies also reside within the CNS, in layer V of the cortex, and hence are not protected from being directly involved by focal MS pathology.
The sensory system has shorter axons, compared to the motor system, and according to the length-dependent axonopathy hypothesis this may protect it. In comparison, motor neurones are much longer than sensory neurones and hence more likely to acquire multiple hits that lead to more rapid or earlier neurodegeneration. The architecture of the sensory system is such that afferent inputs go via first, second and third order neurones that communicate via synapses before reaching the cortex. Each relay axon is relatively short and hence this architecture may provide the sensory system relative protection from MS-associated neurodegeneration. In fact, the first-order sensory neurones are actually bipolar cells with their cell bodies residing outside the central nervous system in the dorsal, or posterior, root ganglia (DRG). A demyelinating plaque affecting the central axonal process is therefore unlikely to kill the neuron residing in the DRG by retrograde neurodegeneration. We know this because sensory radiculopathies that destroy the central neuronal processes before they enter the spinal cord are characterised by preservation of sensory nerve action potentials (SNAPs), which indicates that the neuronal cell bodies and peripheral axons are intact. The compartmentalisation of 1st-order sensory neuronal cell bodies, outside the CNS, away from the MS pathology, may provide the sensory pathway with a better chance of recovery, via axonal sprouting or axonal regrowth.
In comparison the axons from the pyramidal, or motor, neurons extend from the primary motor cortex to synapse on the anterior horn cells, or their equivalents, in the lateral columns of the spinal cord or brain stem respectively. Importantly, there are no motor interneurons, which is why the motor neurones are the longest neurones in the central nervous system. The cell bodies also reside within the CNS, in layer V of the cortex, and hence are not protected from being directly involved by focal MS pathology.
 |
| The motor fibre tracts |
{kind=link}
3. Could reserve capacity explain the different prognostic significance of sensory and motor attacks?
Extending the arguments above that the sensory system may be more sensitive to small lesions due the reporting of percepts, in particular positive phenomena, that as it degenerates, loses neurones and axons, the likelihood of new lesions causing symptoms, in particular positive symptoms, diminishes. In comparison, as the motor system degenerates and compensatory mechanism fail new lesions are more likely to become clinically apparent. If reserve capacity in a particular pathway determines its sensitivity to cause relapses then one could hypothesise that proportion of positive sensory attacks (positive sensory symptoms) will diminish, in comparison to negative sensory attacks (loss of sensation). The latter hypothesis is testable and could be explored in existing databases or prospectively in clinical cohorts."
"Apologies for this post being so technical; I simply want to get the idea into the open to stimulate discussion and debate."
Kalincik et al. Risk of relapse phenotype recurrence in multiple sclerosis. Mult Scler. 2014 Oct;20(11):1511-22.
Objectives: The aim was to analyse risk of relapse phenotype recurrence in multiple sclerosis and to characterise the effect of demographic and clinical features on this phenotype.
Methods: Information about relapses was collected using MSBase, an international observational registry. Associations between relapse phenotypes and history of similar relapses or patient characteristics were tested with multivariable logistic regression models. Tendency of relapse phenotypes to recur sequentially was assessed with principal component analysis.
Results: Among 14,969 eligible patients (89,949 patient-years), 49,279 phenotypically characterised relapses were recorded. Visual and brainstem relapses occurred more frequently in early disease and in younger patients. Sensory relapses were more frequent in early or non-progressive disease. Pyramidal, sphincter and cerebellar relapses were more common in older patients and in progressive disease. Women presented more often with sensory or visual symptoms. Men were more prone to pyramidal, brainstem and cerebellar relapses. Importantly, relapse phenotype was predicted by the phenotypes of previous relapses. (OR = 1.8–5, p = 10-14). Sensory, visual and brainstem relapses showed better recovery than other relapse phenotypes. Relapse severity increased and the ability to recover decreased with age or more advanced disease.
Kalincik et al. Risk of relapse phenotype recurrence in multiple sclerosis. Mult Scler. 2014 Oct;20(11):1511-22.
Objectives: The aim was to analyse risk of relapse phenotype recurrence in multiple sclerosis and to characterise the effect of demographic and clinical features on this phenotype.
Methods: Information about relapses was collected using MSBase, an international observational registry. Associations between relapse phenotypes and history of similar relapses or patient characteristics were tested with multivariable logistic regression models. Tendency of relapse phenotypes to recur sequentially was assessed with principal component analysis.
Results: Among 14,969 eligible patients (89,949 patient-years), 49,279 phenotypically characterised relapses were recorded. Visual and brainstem relapses occurred more frequently in early disease and in younger patients. Sensory relapses were more frequent in early or non-progressive disease. Pyramidal, sphincter and cerebellar relapses were more common in older patients and in progressive disease. Women presented more often with sensory or visual symptoms. Men were more prone to pyramidal, brainstem and cerebellar relapses. Importantly, relapse phenotype was predicted by the phenotypes of previous relapses. (OR = 1.8–5, p = 10-14). Sensory, visual and brainstem relapses showed better recovery than other relapse phenotypes. Relapse severity increased and the ability to recover decreased with age or more advanced disease.
Conclusion: Relapse phenotype was associated with demographic and clinical characteristics, with phenotypic recurrence significantly more common than expected by chance.